Izvedaba cestarine poput receptora. NOD proteini
TLR5 (1q33) izražena na stanicama slezene i leukocita periferne krvi, endotelne stanice. TLR5 prepoznaje flagellin Gram-pozitivnih i Gram-negativnih bakterija te potiče indukciju signalnih kaskada uključuju kemokine, citokine i kostimulacijske molekule.
TLR6 (4p) izražen na leukocita periferne krvi i stanica slezene formira dimere sa TLR2 i prepoznaje peptidoglikan zimosanom.
TLR7 (Hr22.3) eksprimira se u stanicama pluća, placente, slezene i leukocita periferne krvi. Prirodni ligand za TLR7 još nije pronađen. Rad obavlja na TLR7-deficijentnih miševa pokazala da TLR7 prepoznaje imidazola, kao što su R848, koji imaju antivirusna svojstva i sistemska primjena TLR7 agonist daje antivirusni učinak.
TLR8 (Hr22) identificiran s TLR7 i TLR9, izražena u velikim količinama u leukocita periferne krvi i stanica pluća.
Da bi se dobio uvid u funkcionalnu ulogu TLR7 i TLR8 i aktivirati repertoar urođenih imunih stanica, skupina KA Gorden korištenih selektivnih agonista tih receptora. TLR7 agonisti direktno aktivirati pročišćene plasmacytoid DB, te u manjoj mjeri monocita. S druge strane, TLR8-agonisti povećava aktivaciju mijeloidne DC i monocita. Prema tome, TLR7-agonisti su učinkovitiji u usporedbi s TLR8-agonista koji inducira IFN-a i IFN-regulatornih citokina (IFN-inducibilni protein i IFN inducirati T-stanica kemoatraktant MLPK čovjeka). Agonisti TLR8 izazvanih proupalnih citokina i kemokina (TNF-a, IL-12, i M1R-1a).
Međutim, prirodni ligand za TLR8 To se još nije otkriven, na sličan način osjetljivi na TLR7 imidazola spojeve s antivirusnim djelovanjem. Također je otkrio da TLR8 igra ključnu ulogu u prepoznavanju pojedinačnih RNA virusa (Coxsackie B i parechovirusa 1).

TLR9 (Zr21.3) lokalizirana unutar stanice u endoplazmatskom retikulumu, i nakon toga izmiještanjem nakon stimulacije na endo soma TLR9 uključena u prepoznavanje nemetiliranih CpG-motiva bakterijske DNA, koja je dokazano u eksperimentima s miševima i staničnih linija deficijentnih u TLR9. Povlačenja svih CpG induciranih učinaka (produkciju citokina, kostimulatorne molekule-ing, proliferacije limfocita).
TLR10-23 (4r14) otvorena je nedavno. Međutim, njihove funkcije i ligandi dalje dosad nepoznate. To je otkrila da su pretežno su izraženi u stanice imunološkog sustava u većoj mjeri na B limfocite i DC plazmotsitoidnyh. Smatra se da je TLR10 i druge Toll-like receptori uključeni u imunološki odgovor, a može djelovati kao ko-receptora, kao TLR1 i TLR6.
Cestarine poput receptori prepoznaju konzervirane molekularni proizvodi od različitih klasa patogena, uključujući gram-pozitivne i gram-negativne bakterije, DNA i RNA viruse, gljivice i protozoe. Priznanje TLRs ligandi dovodi do razvoja niza signala koji uzrokuje brzo imunološki odgovor potrebno za ubijanje patogena. Signali koje primaju TLRs, također su odgovorni za izazivanje DC sazrijevanja i naknadne indukcije adaptivnog imunološkog odgovora.
NOD proteini. Nedavna istraživanja su pokazala postojanje drugih imunoloških urođene prepoznavanje receptora, takozvani nukleotid-vezujuće domene oligomerizing (nukleotid-vezujući oligomerizacijsku domenu - NOD) proteina koji djeluju u citoplazmi stanica domaćina. NOD receptori su uključeni u prepoznavanju bakterijskog peptidoglikana intracelularni [263, 640].
NOD2 prepoznaje konzervirana muramilpeptidnye stanica struktura stijenke svih bakterija, a NOD1 prepoznaje je prisutan u staničnom zidu gram-negativnih bakterija peptidoglikan diaminopimelinska kiselina. Kao TLRs, funkcionalni značaj NOD signala identificirana kod pacijenata s NOD-mutacija koje su podložniji kroničnim bolestima bakterijskog podrijetla.
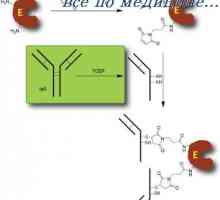 Aktivacija model u stanicama. indukciju antitijela
Aktivacija model u stanicama. indukciju antitijela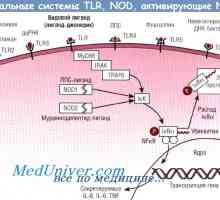 Izvedbe obrazraspoznayuschih PRRS receptore. Cestarine poput receptori
Izvedbe obrazraspoznayuschih PRRS receptore. Cestarine poput receptori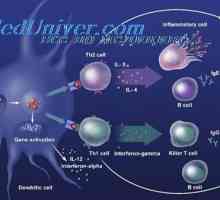 Proteini toplinskog šoka (hsp). Učinci imunog odgovora na hsp
Proteini toplinskog šoka (hsp). Učinci imunog odgovora na hsp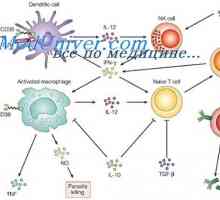 Ligandi receptora efektora prirođenog imuniteta. Peptidoglikanski, lipopeptidi
Ligandi receptora efektora prirođenog imuniteta. Peptidoglikanski, lipopeptidi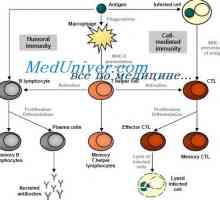 Signali cestarine slične receptore. Uloga molekule MyD88
Signali cestarine slične receptore. Uloga molekule MyD88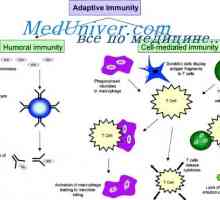 Limfnog ili plasmacytoid dendritične stanice. Funkcija dendritičkih stanica
Limfnog ili plasmacytoid dendritične stanice. Funkcija dendritičkih stanica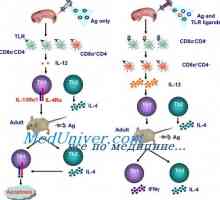 Akcija aktivacija imunomodulatori u dendritičke stanice. Sazrijevanje dendritičke stanice
Akcija aktivacija imunomodulatori u dendritičke stanice. Sazrijevanje dendritičke stanice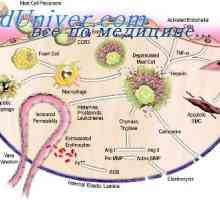 Interakcija T-helper stanice s dendritičnim stanicama. Diferencijacija stanica CD4
Interakcija T-helper stanice s dendritičnim stanicama. Diferencijacija stanica CD4 Citokina razine aktivacije dendritičkih stanica. Proizvodnja citokina pomoću dendritičke stanice…
Citokina razine aktivacije dendritičkih stanica. Proizvodnja citokina pomoću dendritičke stanice… Citotoksičnost prirodnih stanica ubojica. Učinak imunomodulatora na NK stanicama
Citotoksičnost prirodnih stanica ubojica. Učinak imunomodulatora na NK stanicama Učinak Immunovac-sn-4 Mononuklearne stanice. Imunofenotip leukociti ispod imunomodulatori
Učinak Immunovac-sn-4 Mononuklearne stanice. Imunofenotip leukociti ispod imunomodulatori Utjecaj imunomodulatori na citokina. proizvodnja citokina nakon stimulacije stanica slezene
Utjecaj imunomodulatori na citokina. proizvodnja citokina nakon stimulacije stanica slezene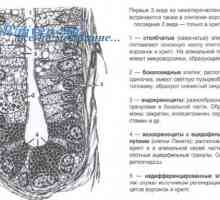 Antimikrobni peptidi crijeva funkcije, poslove u imunološkom sustavu
Antimikrobni peptidi crijeva funkcije, poslove u imunološkom sustavu- Epitelne bakterije kontrole u crijevima: Mamre i PRR
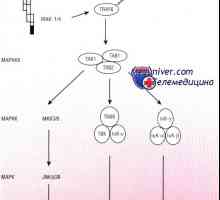 Imunološki odgovor na intestinalni epitel i bakterije aktivaciju put apoptoze
Imunološki odgovor na intestinalni epitel i bakterije aktivaciju put apoptoze- Interakcija s crijevne mikroflore i njihov međusobni utjecaj
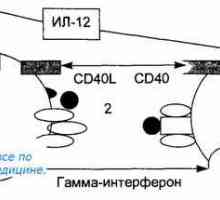 Prezentacija antigena. prepoznavanje antigena. Interakcija T-pomoćničkih (Th1) sa stanicama s…
Prezentacija antigena. prepoznavanje antigena. Interakcija T-pomoćničkih (Th1) sa stanicama s…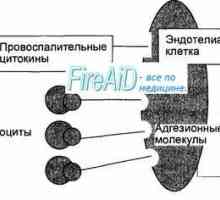 Korak imunološki odgovor. Oblici imunološkog odgovora. Upala. Rano zaštitni upalni odgovor.
Korak imunološki odgovor. Oblici imunološkog odgovora. Upala. Rano zaštitni upalni odgovor. Veliki Medicinska Enciklopedija IC nevronet. lijekovi
Veliki Medicinska Enciklopedija IC nevronet. lijekovi- Stanični protein TLR5 - ključ za liječenje reumatoidnog artritisa
 Leukemija, simptomi, liječenje, uzroci
Leukemija, simptomi, liječenje, uzroci
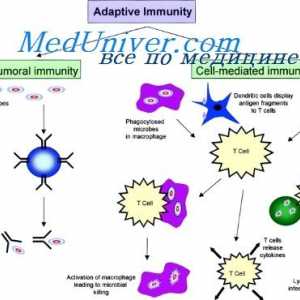 Limfnog ili plasmacytoid dendritične stanice. Funkcija dendritičkih stanica
Limfnog ili plasmacytoid dendritične stanice. Funkcija dendritičkih stanica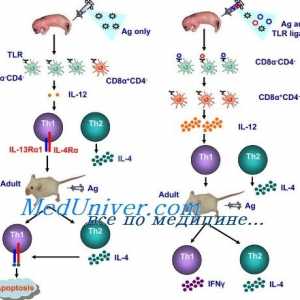 Akcija aktivacija imunomodulatori u dendritičke stanice. Sazrijevanje dendritičke stanice
Akcija aktivacija imunomodulatori u dendritičke stanice. Sazrijevanje dendritičke stanice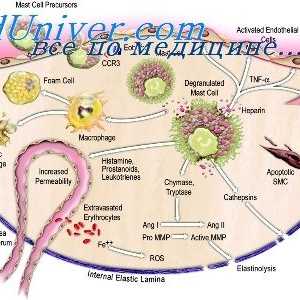 Interakcija T-helper stanice s dendritičnim stanicama. Diferencijacija stanica CD4
Interakcija T-helper stanice s dendritičnim stanicama. Diferencijacija stanica CD4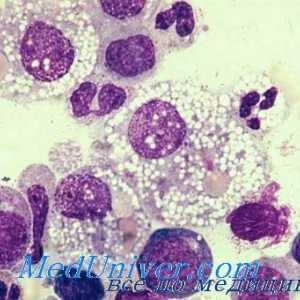 Utjecaj imunomodulatori na citokina. proizvodnja citokina nakon stimulacije stanica slezene
Utjecaj imunomodulatori na citokina. proizvodnja citokina nakon stimulacije stanica slezene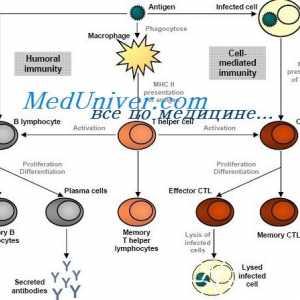 Signali cestarine slične receptore. Uloga molekule MyD88
Signali cestarine slične receptore. Uloga molekule MyD88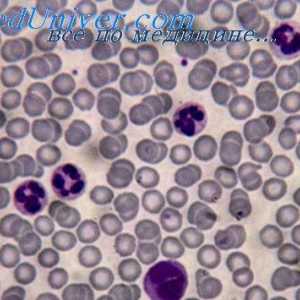 Citokina razine aktivacije dendritičkih stanica. Proizvodnja citokina pomoću dendritičke stanice…
Citokina razine aktivacije dendritičkih stanica. Proizvodnja citokina pomoću dendritičke stanice…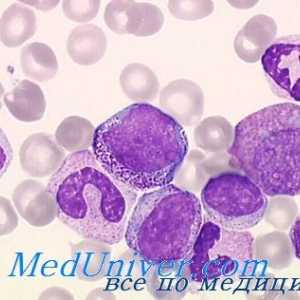 Učinak Immunovac-sn-4 Mononuklearne stanice. Imunofenotip leukociti ispod imunomodulatori
Učinak Immunovac-sn-4 Mononuklearne stanice. Imunofenotip leukociti ispod imunomodulatori Proteini toplinskog šoka (hsp). Učinci imunog odgovora na hsp
Proteini toplinskog šoka (hsp). Učinci imunog odgovora na hsp