Onkologiya-
B.P.Kopnin
Rak ruski istraživački centar. Blokhin RAMS, Moskva
izvor RosOncoWeb.Ru
01 0203 04Uvod.
Karcinogeneza - Stupnjeviti proces andother mutacija akumulacije genetskih promjena koje vode do poremećaja klyuchevyhkletochnyh funkcijama kao što su regulacije proliferacije i diferencijacije, prirodni staničnu smrt (apoptozu) morfogenetskog reaktsiykletki, a vjerojatno i neučinkovitog rada faktorovspetsificheskogo i nespecifičnih antitumorskoj immuniteta.Tolko kombinaciji kao promjene stečena, najčešće kao rezultat prilično duge evolucije maligne klonova, u kojem Tu je izbor stanica sa željenim značajkama može osigurati razvoj malignih bolesti. Veroyatnostvozniknoveniya u istoj ćeliji više genetski izmeneniyrezko povećava s poremećajima sustava kontroliruyuschihtselostnost genoma. Stoga, mutacije koje dovode do genetske nestabilnosti, također su bitan korak u progresiji tumora. Bolesna, neke kongenitalne anomalije sustava genetski kontrolyayavlyayutsya čimbenik određivanja neizbježnu pojavu novih formacija: oni su tako povećati vjerojatnost pojave svake kletkeorganizma različitih onkogena mutacija koje pojedinac ransheili kasnije u nekim stanicama množe klonove pod davleniemotbora nužno akumulirati potrebnu skup izmeneniyi tumora obrasca.
Značajan napredak u razumijevanju mehanizama karcinogeneze svyazans prvog otvaranja i protonkogenov onkogena, a onda - opuholevyhsupressorov i mutatorskima gene. Onkogeni - stanica ili virus (virus uvedeni u stanicu) gena čija ekspresija može privestik razvoj tumora. Proto-onkogena - normalna kletochnyegeny, poboljšati ili izmijeniti funkcije od kojih ih vonkogeny čini. Tumor supresor (antioncogene, recesivna opuholevyegeny) - staničnih gena čiji inaktivacija oštro uvelichivaetveroyatnost pojave tumora i oporavka funkcije, s druge strane, mogu spriječiti rast tumorskih stanica. Treba napomenuti, da je iz tzv tumora potiskivača "mutatorski"geni koji geni disfunkcija koja na neki sposobomuvelichivaet tempo mutacija i / ili druge geneticheskihizmeneny, ne može utjecati na rast tumorskih stanica. Odnakoih inaktivacije tako uvelike povećava vjerojatnost poyavleniyarazlichnyh onkogena mutacija koje formiranje tumora stanovitsyalish pitanje vremena.
Pripadaju onkogena ili tumorske potiskivača opredelyaetsyaneskolkimi kriterija: a) prirodni znak mijenja strukturyi / ili ekspresije gena u stanicama ili nekom razlichnyhnovoobrazovaniy- b) izgled mladih mlađe vozrasteopredelennyh oblika tumora kod pojedinaca s prenosi nasledstvugerminalnymi (tj došlo u seksualni stanica) mutatsiyamidannogo gena- c) oštar porast u učestalosti pojave opuholeyu transgeničnih životinja koje eksprimiraju genski aktiviranog formudannogo - u slučaju onkogena ili ležajeva (inaktiviruyuschiemutatsii"nokaut") Gen - u slučaju opuholevyhsupressorov- g) sposobnost da induciraju u kultiviranim vitrokletkah u morfološkom transformacije i / ili neograničenog rasta (onkogeni) ili suzbijanje rasta stanica i / ili transformacije (vyrazhennostipriznakov tumor supresor).
Posljednja dva desetljeća obilježena brzim otkrytiemvse više onkogeni i tumor prigušivači. Do nastoyaschemuvremeni zna o stotinama potencijalnih onkogena (kletochnyhi virus) i oko dvadesetak tumor supresorskih. Byliopisany genetski događaja koji vode do aktivacije protoonkogenovili inaktivacija tumor supresor [1-6]. Nađeno chtomehanizm djelovanje virusne onkogene povezanih s aktivacijskim kletochnyhprotoonkogenov (retrovirusi) ili inaktivacije tumora potiskivača (DNA virusa) [7-11]. Karakteristična za oblike iliinyh ljudskih tumora i promjena opuholevyhsupressorov onkogena, uključujući visoko specifičan poremećaj dijagnoza ispolzuemyedlya [3.12] (Tablica, 1, 2).
Tablica 1.
Neke promjene u proto-onkogena obilježje novoobrazovaniycheloveka
| protooncogen | funkcija proteina | promjene | neoplazme * |
|---|---|---|---|
| RET (GDNF-R) | receptor tirozin kinaze | a) aktiviranje točkastom mutacijom-vodilice; b) rekombinacije ob tvorbu himerne gene Ret / PTC, šifrirati vrelišta trajno acti-receptora virovanny | sindroma multiple endokrine neop-lijeni (MEN2a, MEN2b), medularni i papilarnog karcinoma štitnjače * |
| ERBB1 (EGF-R) | receptor tirozin kinaze | Pojačanje i prekomjernom ekspresijom gena | glioblastoma i drugih tumora neurogene |
| ErbB2 (HER2) | receptor tirozin kinaze | Amplifikacija i / ili prekomjerna ekspresija gena | raka dojke |
| PDGF-Rb | receptor tirozin kinaze | kromosoma-translacijska katsii tvore Chi-dimenzionalni gena TEL / PDGF-Rb, CVE6 / PDGF-Rb, Cody stalno acti-al-receptori virovannye | kronična mijeloidna leukemija, akutne mijeloblastične leukemije |
| SRC | non-receptor tirozin kinaze | mutacije na kodonu 531, ukidanje negativne regulacije kinazne aktivnosti | dio debelog crijeva u kasnim stadijima |
| K-ras, N-ras, H-ras | sudjeluje u prijenosu signalima mitoze i regulacije morfogeneti-cal reakcije | mutacije u kodona 12,13,61, uzrokujući formiranje trajno aktivirani GTP-bound obliku Ras | 60-80% slučajeva raka gušterače zhelezy- 25-30% različitih solidnih tumora i leukemija |
| PRAD1 / tsiklinD1 | regulira ljepilo-ciklus točan | Amplifikacija i / ili prekomjerna ekspresija gena | rak dojke i žlijezde slinovnice |
| C-myc | TransKom faktor RTOC, regulatori liruet kletoch-og ciklusa i djelovanje telomeraze | a) trans-kromosom položaj, prijenos gena pod kontrolom regulatornog gena imunoglobulina regulacijskih elemenata; b) pojačavanje i / ili prekomjerna ekspresija gena- mutacija stabilizira protein-ing | a) limfom Berkita b) više oblika tumora |
| CTNNB1 (b-katenina) | a) faktor transkripcije-onny, reguliret c-myc i ciklin D1; b) vezanje na kadherina-nastavi postoji u put-vania ljepilom vladina kontakata | mutacija koje povećavaju količinu nevezanog-provodljivog-vezan na E-kadherina b-katenina, koji djeluje kao faktor transkripcije | obiteljska adenomatozna polipoza debelog crijeva kishki- različite oblike sporadični |
| BCL2 | inhibira apo ptozu, reguliranjem propusnosti mitohondrija i jezgre-azovite-tion membrane | kromosoma-translacijska katsii, prijenos gena pod kontrolom regulatornog gena imunoglobulina regulacijskih elemenata | folikularni limfom |
| ABL | regulira stanični ciklus i apoptozu | kromosom-translacijske katsii dovodi do Obra-simacije himerni gena BCR / ABL, toryh-proizvoda stimuliraju proliferaciju stanica i apoptozu podvlyayut | sve kronična mijeloidna leukemija, akutna limfatična leukemija dijelom-blastic |
| MDM2 | deaktivira p53 i PRB | Amplifikacija i / ili prekomjerna ekspresija gena | dio osteosarkoma i sarkomi mekanog tkiva |
Tablica 2.
Oblike humanih tumora koji proizlaze iz inaktivacije nekotoryhopuholevyh potiskivača i mutatorski gena
| gen | funkcija proteina | neoplazme * |
|---|---|---|
| p53 | transkripcijski faktor-liruet regulatori staničnog ciklusa i apoptoze, prati integritet genoma | Li-Fraumeni sindrom i većina oblika sporadični |
| INK4a-ARF | Inhibicija Cdk4 ** ** aktivacija p53 | nasljedna melanom i mnogi sporadični tumori |
| Rb | kontrolira ulazak u S-fazu, podešavanje aktivnost transkripcijski faktor E2F | nasljedna retinoblastom i mnogi oblici sporadičnih tumora |
| TBR-II | receptor za druge vrste citokina TGF-b | nasljedne i sporadični raka debelog crijeva |
| Smad2, 3 Smad | odašiljanje signala iz aktiviranih TGF-b receptora za Smad4 | raka debelog crijeva, pluća, gušterače |
| SMAD4 / DPC4 | transkripcijski faktor-OPOS-reduet učinak citokina TGF-b, što dovodi do aktivacije ingibito-rova CDK - p21WAF1, p27KIP1, p15INK4b | juvenilni polyposis gamartomatoz-og kishechnika- želudac i različiti oblici sporadični |
| E-kadherina | uključeni u interakcije stanica-stanica, pokreće deystviyah- prijenos signala koji aktiviraju p53, p27Kip1 | nasljedni oblici raka želuca i mnogi oblici sporadični |
| APC | veže i uništava tsitoplaz nematik-b-katenina, sprečava nastajanje kompleksa-transkripcijski b-katenina / TCF | porodične adenom-tozny polipoza i sporadična tumora debelog crijeva |
| VHL | inhibira ekspresiju VEGF gena (faktor rasta vaskularnog endotela) i drugih gena koji se aktivira tijekom hipoksije | sindrom von Hippel Lindau (višestruki hemangiomi) - jasno stanica karcinoma bubrežnih stanica |
| WT1 | transkripcijski faktor koji vežu na p53 modulira donose gene Ex-Press p53 reagiraju | nasljedni nefroblas jem (Wilmsov tumor) |
| PTEN / MMAC1 | fosfataza- potiče apoptozu inhibicije aktivnosti PI3K-PKB / Akt signalnog puta | Cowden bolest (set-vladina hamartoma) - mnogi tumori sporadične |
| NF1 (neyrofibromin) | Obitelj protein-GAP ras onkogena transformacija iz aktivnog u neaktivnom obliku | Prvi tip neurofibromatoza |
| NF2 (Merlin) | uključeni u interakciju s membranom na stanični skelet | neurofibromatoza drugog tipa-sporadično meningitis Ohma, mezotelioma i sur. Tumor |
| BRCA1 | povećava aktivnost p53 i drugim faktorima transkripcije, vezanje na RAD51 sudjeluje u učenju i / ili popravak DNA | nasljedni karcinom dojke i različiti oblici yaichnikov- sporadične tumora |
| BRCA2 | faktor traskriptsionny s aktivnostima histon acetil transferazy- vezivanje za RAD51 je uključen u popravak DNA | nasljedni karcinom dojke i različiti oblici yaichnikov- sporadične tumora |
| MSH2, MLH1, PMS1, PMS2 | pojedinačna popravak DNA stranice (mismatch popravak) | nonpolyposis raka debelog crijeva i yaichnikov- više sporadičnih tumora |
** mjesto INK4a / ARF kodira dva proteina: p16INK4a - tsiklinzavisimyhkinaz inhibitor cdk4 / 6 i p19ARF (Alternativni Reading Frame) - alternativnoyramki čitanje produkt koji se veže na p53 i MDM2, sprječavanja interakciju sprječava degradaciju p53 [13, 14]. Delecije i mnogo tochechnyemutatsii mjesto INK4a / ARF supressornyhaktivnostey uzrokuje inaktivaciju istovremeno oba proteina [15].
Međutim, dugo vremena znanje o svakoj od onkogena ili opuholevyhsupressorov izvedeni u zasebnim, uglavnom nepovezane. I samo u posljednjih nekoliko godina postalo je vyrisovyvatsyaobschaya slika, što pokazuje da je velika većina izvestnyhprotoonkogenov i tumor prigušivači su komponentamineskolkih zajedničke signalni putevi koji kontroliraju stanični ciklus, apoptoza, integritet genoma i morfogeneze reakcije differentsirovkukletok. Očito, promjene u tim signalnim putevima u kontsekontsov i dovesti do razvoja malignog novoobrazovaniy.Dalee pruža informacije o glavnim ciljevima djelovanja onkogenovi tumora smetnji.
1. Onkogeni i tumorski supresori u regulaciji staničnog ciklusaOsnova stvaranja tumora je prekomjerno razmnožavanje opredelennyhkletok. To je samo prirodna, dakle, da prekršaji regulyatsiikletochnogo ciklus je sastavni i temeljna priznakomneoplasticheskoy stanice. "motor" staničnog ciklusa, kao što je poznato, u slijedu su uzastopna drugdruga Aktivnost kinaza ovisnih o ciklinu [16] (Sl. 1). Svaki tsiklinzavisimayakinaza (SDK) je katalitička podjedinica holofermentnogokompleksa za koje djelovanje zahtijeva prisutnost aktivatornoysubedinitsy - ciklin. Regulacija rezultat aktivnosti SDK osuschestvlyaetsyaza režiji razini promijeniti određene ciklina vopredelennye fazi staničnog ciklusa. Nadalje, aktivnost mijenja Sdkreguliruetsya fosforilacije određenog aminokislotnyhostatkov. Aktivni oblik ciklin-CDK kompleksi fosforiliruyutregulyatornye proteine koji kontroliraju protok faze.

Sl. 1. Kretanje staničnog ciklusa određena posledovatelnoyaktivatsiey različitih ciklin - Cdk. Većina njih je učinak aktiviranja meta zabranjivanje onkogena ili supresorskih deystviyaopuholevyh
Utvrđeno je da je učinak mnogo proto-onkogena i tumor supressorovnapravleno regulaciju različitih ciklin kompleksa - Cdk.Belkovye proizvodi većina njih povećati aktivnost tsiklinzavisimyhkinaz odgovoran za početne faze predmitotičke fazyG1 (kompleksi ciklin D1 - D3 c Sdk4 ili Sdk6 ovisno ottipa stanica) prijelaz iz G1 faze u sintezi DNA (ciklin E - Sdk2) (Slika 1.). Nadalje, neki proto-onkogena i tumor supressoryreguliruyut aktivnost ciklin A - Sdk2 (dlyareplikatsii potrebno DNA) i ciklin B - CDK1 (također poznat kao Cdc2 CDK1 -, je potrebno za prijelaz iz G2 u mitozi).
Primarni supstrat kompleksa ciklin D - Cdk4 i ciklin D - Cdk6yavlyaetsya Tumor supresor pRb i Rb kao što su proteini i-P105 r130.pRb defosforilirana te njegovi homolozi koje se ne dijele i stanica vproliferiruyuschih stanice u ranoj fazi G1-[17]. VTAK stanje vežu i blok transkripcije kompleksyE2F - DP (E2F-1, -2, -3, -4, -5 i DP-1, -2, -3), aktivnostryada reguliranje gena čiji proizvodi su potrebne za početak i prohozhdeniyaS- faza. Posebno, E2F-DP reguliraju ekspresiju gena za timidin kinaze, dihidrofolat reduktaza, ciklina E, ciklin A, PCNA, DNA polimerazya et al. [18]. Vezanje proteina obitelji E2F c PRB ingibiruetih transkripcijsku aktivnost. Kada mitogenih signala vyzyvaemyhrostovymi faktori pRb sredinom G1 fazi fosforilirani kompleksomtsiklin D - (Sdk4 ili ciklin D - CDK6), što uzrokuje vysvobozhdenietranskriptsionnyh E2F čimbenika - DP iz kompleksa s pRb i njegovu aktivaciju [17]. Jedna posljedica toga je poticanje transkriptsiigena ciklin E, pri čemu su aktivirani kompleksi ciklin - Cdk2 i fosforilaciju URŽ-a. Tako voznikaetregulyatornaya zglob potporni aktivnost transkriptsionnyhfaktorov E2F - DP i kontrolirani geni pružanja replikatsiyuDNK (vidi Sl. 2.). Nakon završetka S-faze postaje pRb defosforilirovannoesostoyanie u kojoj blokira aktivnost E2F - DP i nakon unosa S-faze (za njegovo pokretanje zahtijeva novi kompleksi aktiviraju mitogennyystimul ciklin D - Cdk4,6). Tako, pRb tumor supresor igra ključnu ulogu u regulaciju vhozhdeniyakletki u S-fazu.
Produkti proto-onkogena mnogih su signalnyhputey komponente koje su odgovorne za aktivaciju ciklina D - CDK4 (6), i ciklin E - CDK2 odgovor na faktore rasta i / iliadgeziyu stanica na izvanstaničnog matriksa proteina (slika 2.). Tako je vezanje receptora faktora rasta s njihovim ligandima indutsiruetdimerizatsiyu autofosforilacija i receptor (jedan subedinitsadimera fosforilira druge tirozin). To je, pak, uzrokuje interakcija receptor tirozin kinaze s mnogo signalnymibelkami sadrži SH2-vezne domene, fosfotirozin. Na primjer, aktivirani receptori faktora rasta trombocita (PDGF-R '') u interakciji s SH2 domenama-proteina, kao što fosfatidilinozitol-3`-kinaze (PI3K), fosfolipaze C (PLC), latentni -g1 oblika transkriptsionnyhfaktorov STAT proteina i adapter Grb2, odašiljanja signal belkamRas [19-21]. Vezanje svaki od tih proteina s preklapanjem fosfotirozinamiretseptora uzrokuje aktivaciju signalnih puteva, a zatim se vrši aktivaciju u jezgri transkripcije kita faktorovi ekspresiju specifičnih gena (Sl. 2). Posebno indutsiruemyyGrb2 prijelaz na aktivirani Ras protein (GTP-vezan) sostoyanievedet stimulirati neke od njegovih efektora, uključujući serin treoninovyhkinaz Raf * i MEKK, aktivira MAP (mitogenima aktivirana proteinska kinaza kaskada) [20,22]. Krajnji produkti tih kaskada, ERK (MAPK), p38 i JNK (SAPK), premještena iz citoplazme u jezgru gdje se fosforilira i aktivira brojne podloge, tomchisle takvih transkripcijskih faktora kao što su, Elk1, Ets1 *, Ets2 *, Jun *, ATF2 , TCF i dr. To je, pak, uzrokuje aktivatsiyuryada druge faktore transkripcije. Tako, Elk1 tvoreći komplekss SRF (serum Response Factor), započinje transkripciju gena koji sadrže u svojim SRE promotor elemenata, kao što FOS * gena.

Sl. 2. Proizvodi mnogih protoonkogena i tumora supressorovreguliruyut aktivnošću o ciklinu ovisnih kinaza za fosforiliranje pRb.Fosforilirovanie PRB, kao i njegove vežu na sljedeći viralnom onkoproteinima uzrokujući oslobađanje i aktivaciju transkripcijskih kompleksovE2F - DP, povećanje ekspresije gena čiji su proizvodi neobhodimydlya prolaz S-faze
Slične reakcije su promatrane nakon vezanja integrina (receptor posreduje adheziju stanica) s proteinima ekstracelularnog matriksa.Takoe interakcije uzrokuje aktiviranje i autofosforilirovaniekinazy FAK (Fokalna Adhezijska Kinaza), pri čemu se svyazyvaetsyas SH2 domena protoonkobelka Src, što zauzvrat vyzyvaetrekrutirovanie Grb2 protein adapter, aktivacija Ras i MAP kinaznyhkaskadov (Slika 2).
Posljedica promjena u aktivnosti brojnih faktora transkripcije inducirana aktivacija MAP kinaza je povećanje ekspressiigena ciklin D1 (pretpostavlja se da je odgovoran za belkiJun, Ets1, Ets2 [21]). Osim toga, mitogene signale povyshayutekspressiyu myc, koji uzrokuje povećanje aktivnosti tsiklinzavisimyhkinaz djeluju u G1 (ciklin D - cdk4 i ciklin E - Cdk2). Etosvyazano da Myc, prvo, transaktivira gena Cdc25a- fosfatazu koja uklanja inhibitorni fosforilaciju Cdk2 i Cdk4po Thr-14 i Tyr-15, i drugo smanjuje ekspresiju ingibitoraSdk2, p27KIP1a [23-26]. Myc mehanizmi aktivacije pod utjecajem rostovyhfaktorov studirao do loše. Pretpostavlja se da to može privoditkak Ras-neovisne signalne putove aktivirane onkoproteina Src, ili Ras-Raf-MAP-kinaze kaskade uzrokuje aktivaciju Ets1i / ili E2F (myc gena obuhvaća elemente koji odgovaraju promotor dlyaetih transkripcijskih faktora [22,26]) ,
Mnogi sudionici signalnih putova koji posreduju deystvierostovyh kao odgovor na aktivaciju ciklin-ovisne kinaze faktora i, dakle, poticanje stanične diobe, su proto-onkogena. Izmeneniyaih struktura (mutacije), koji je rezultat bijega vozdeystviyanegativnyh regulacijski faktori i / ili trajno povysheniyuekspressii prevede u protooncogenes onkogena takve [1,3,5] .The proizvod identificiran onkogeni su sve etaže regulyatsiimitogennogo signala [5]: čimbenici rasta - PDGF-b (SIS), FGF1i drugi receptor tirozin kinaze - EGF-R (ErbB), HGF-R (Met), i drugi retencije obitelj proteina Ras - K-Ras, H-Ras i N-Ras- effektoryRas - serin treonin kinaze Raf i Mos- faktori transkripcije-jun, Ets1, Myc itd-i, na kraju, ciklin D1 (Prad1). Skladyvaetsyavpechatlenie da detaljna analiza svake novoobrazovaniivyyavlyayutsya promjene barem jednog od komponenata (signalnyhputey protooncogenes) uzrokuje trajnu stimulaciju aktivnostitsiklinzavisimyh kinaze i pokretanje stanične diobe je zavisimostiot djelovanje faktora rasta.
Zanimljivo je signalni put Cdk-Rb-E2F kontrolira ne tolkopRB, ali i mnoge druge supresijskog proteina (Sl. 2). Nekotoryeiz njih inhibitorne podjedinice Cdk (CKIs - Cdk inhibitori), posreduju u zaustavljanje staničnog ciklusa u odgovor na razlichnyevnekletochnye i unutarstanične signale [16]. Obitelj Identifitsirovanodva CKIs: INK 4 i Cip / Kip. Bivši sastoji se od četiri člana, uključujući i tumor suppressors p15INK4b i p16INK4a antitijelom. Proteini Ink4obladayut dovoljno uzak specifičnost: združuju Cdk4 i CDK6, sprečavaju formiranje njihovih kompleksa s ciklinima D [16,27] .Semeystvo Cip / Kip se sastoji od tri dijela: p21WAF1 / CIP1, p57KIP2 p27KIP1ai. Ovi proteini se vežu na i inhibira ciklin već u potpunosti sformirovannyekompleksy D - CDK4 (6), ciklin E - Cdk2 i ciklin A - Cdk2.Krome dodavanja, p21WAF1 / CIP1 je sposoban za blokiranje i složenih tsiklinB - Cdc2 je odgovoran za promociju G2-faze stupanja mitoza [16,27]. I p21WAF1 / CIP1 i p27KIP1a posredovati utjecaj drugihsupressornyh proteina. p21WAF1 / CIP1 je glavno djelovanje misheneytransaktivatsionnogo p53, i time odvodnik uključen u regulaciju aktivnosti stabilnost / p53 (p19ARF, ATM, WT1 [13,14,28,29] ili njegova transkripcijsku aktivnost (BRCA1 ip33ING1 [30-32 ]), (BRCA1 i WT1 sposobna aktivirati p21WAF1 / CIP1takzhe i još nepoznatih mehanizama p53 neovisno [31,33]).
Uz r15INK4b, p27KIP1a je ključna komponenta peredachiingibitornyh signali inducirana vezanjem TGF-b sa svoimiretseptorami (Sl. 2). Nedavno je nađeno da aktivira retseptoryTGF-b fosforilira specifične signalizaciju djelovatelji belkiSmad2 i Smad3, uzrokujući njihovo vezivanje za tumora supressoromSmad4. Nastali kompleks vyadro pomjeriti iz citoplazme, gdje reguliraju transkripciju specifičnih gena u chastnostiingibitorov CDK. Kao rezultat toga, aktivni i p21WAF1 / CIP1, a r15INK4b [34-37]. Zadnje p27KIP1a pomiče iz kompleksa Cdk4 s / 6 formiranje ipodavlyaet njihovih kompleksa s ciklina D, neobhodimyhdlya napredovanje G1 (Sl. 1). Oslobođeni p27KIP1a u svoyuochered veže i inhibira kompleksa ciklin E - Sdk2, otvetstvennyeza početak S-faze. Povećana ekspresija p21WAF1 / CIP1 vedetk također suprimiraju aktivnost ciklina D - Cdk4,6 i tsiklinE - Cdk2. Kao posljedica toga, stanice se zaustavlja u G0 / G1 i S-faze vhoditv (sl. 3).

Sl. 3. Vezanje TGF-b na njegov receptor dovodi obrazovanietranskriptsionnyh kompleksa Smad4 - Smad2,3 koji translotsiruyutsyaiz citoplazme u jezgru. To dovodi do aktivacije niza ciljeva, uključujući pogrešno i ciklin ovisne inhibitor kinaze p21WAF1 / CIP1, p15INK4b, p27KIP1a, što uzrokuje supresiju djelovanja Cdk4,6 i cdk2, otvetstvennyhza napredovanje G1 i ulazak u S fazu (objašnjenje u tekstu)
Inhibitorni učinak na TGF-b, ili hipereksprimira prevlada onkogenovMYC MDM2 [38]. I ako je radnja povezana s aktivacijom myc razlichnyhCdk povećanjem ekspresije Cdc25A [23] i stimulacije degradatsiip27KIP1a [24], Protein MDM2 dodatno degradaciju p53 [13,14] vyzyvaeti inaktivacija pRb [39], čime se otpušta aktivni transkriptsionnyekompleksy E2F - DP. Prema tome, prekomjerna ekspresija protoonkogenovMYC ili MDM2 i inaktiviranjem mutacije tumora supressorahSmad4, p15INK4b, pRb su jedan zajednički posljedica - stanice uskolzayutot inhibitorni učinak na TGF-b, koji se pojavljuje u jako vazhnymdlya epitelnim tumorima, posebice raka gušterače kishechnikai [40,41] ,
Jedan od najistaknutijih dostignuća u posljednje dvije godine yavilasidentifikatsiya još jedan važan signalni put, vrlo chastonarushayuschegosya u različitim humanim tumorima i provodi je vjerojatno regulacija staničnog ciklusa, ovisno o sostoyaniyamembrannyh pod-membranska struktura i stanicama [42] (vidi. Sl. 4) .Okazalos, da je povezana s E-kadherina stanje b-kateninmozhet djeluju kao transkripcijski faktor. Citoplazma onsvyazyvaetsya drugom transkripcijski faktor Tcf4 nakon chegokompleksy b-katenina - Tcf4 translociraju u jezgru i aktiviruyutgeny koji u svom sastavu specifične vježbanje reagiraju elementy.Odnimi brojne ciljeve transaktivacija djelovanje kompleksab-katenina - Tcf4 se ciklin D1 gena [43] i myc [44]. Opuholevyysupressor APC, prirođene mutacije u koji uzrokuju razvoj crijeva adenomatoznogopolipoza veže slobodne citoplazmatski b-katenina, koja uzrokuje degradaciju potonjeg [45,46]. Tako, poticanje formiranja kompleksa inaktivatsiyaARS b-katenina - Tcf4, povyshaettranskriptsiyu ciklin D1 gena myc i kao posljedica toga, dolazi do aktivatsiitsiklinzavisimyh kinaze odgovorne za poticanje vhodv G1 i S fazu (slika 4). Isti učinak vodi mutatsiib-katenina, koji povećavaju stabilnost u citoplazmi (takiemutatsii pronađena kod pacijenata s obiteljskom polipoza, nisu APC imeyuschihmutatsy [43.47] - većina tih stranica utječe b-katenina, fosforilirani glikogen sintetaze kinaze-3b, GSK 3b). Shodnayakartina također opažena kad se aktiviraju onkogena WNT1 (krila / Int1) (Slika 4). Veže ga Wnt1 produkt (član obitelji cisteina bogatyhglikozilirovannyh signalnih proteina) na njegov receptor (frizzled) uzrokuje citoplazmatski protein DSH do membrane gdje inhibira djelovanje kinaze GSK-3b, koji fosforiliruyab-katenina i APS, potiče vezanje i razgradnja b -katenina.Takim način Wnt1 izazvana inaktivacija GSK-3bprivodit stabilizirati i povećati unutarstanične kontsetratsiitsitoplazmaticheskogo b-katenina, što povećava vjerojatnost transkripcije kompleksa obrazovaniyaaktivnyh s b-katenina s semeystvaTcf / Lef1 faktora [48]. Moguće je da mutacije u E-kadherina takzhemogut biti odgovoran za poticanje signalnih putova oposreduemyhtranskriptsionnymi aktivnosti b-katenina.

Sl. 4. mutacije u tumor supresor APC i beta-katenina, kaki aktivaciju onkogena wnt1, stimuliraju formiranje b-katenina transkriptsionnyhkompleksov - Tcf4, regulatorni geni myc i ciklin D1.V rezultat povećane aktivnosti niza ciklina - Cdk (objašnjenje u tekstu)
Da zaključimo ovaj dio, napominje se da je većina izvestnyhprotoonkogenov i tumora prigušivači na bilo koji način reguliruyutaktivnost ciklin ovisne kinaze odgovorne za ulazak u S-fazukletochnogo ciklusa. Produkti neke stanice (MDM2) ili virusa (virus T antigen SV40, adenovirus E1A, E7 HPV i sur.), I inaktivira onkogenovsvyazyvayut glavni supstrat takav Cdk - pRb. Očigledno, kršeći signalnih puteva > Cdk2,4 / 6 > pRb > E2F / DPyavlyayutsya potrebno za pojavu stalno proliferiruyuschihneoplasticheskih stanica.
 10% Prirođenih srčanih bolesti nisu naslijeđene od roditelja
10% Prirođenih srčanih bolesti nisu naslijeđene od roditelja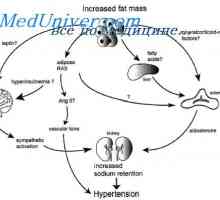 Živaca uzrokuje pretilosti. Genetski uzroci pretilosti
Živaca uzrokuje pretilosti. Genetski uzroci pretilosti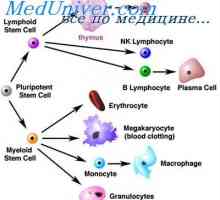 Stanice koje utječu na matičnim stanicama. Generacijskih ciklus matičnih stanica
Stanice koje utječu na matičnim stanicama. Generacijskih ciklus matičnih stanica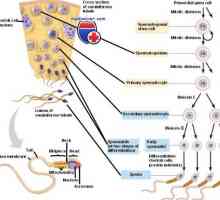 Formiranje sperme. Spermatotsitogenez i spermatogeneza
Formiranje sperme. Spermatotsitogenez i spermatogeneza- Proteini imunološkog sustava uzrokuje DNK mutacije koje dovode do raka
- Matičnih stanica terapija ne dovodi do raka
- Znanstvenici su otkrili genetski uzrok akutne limfoblastične leukemije
- Genetske varijante arterijske hipertenzije
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
 Onkologiya-
Onkologiya-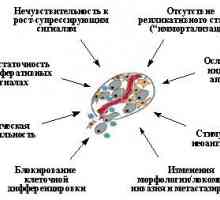 Onkologiya-
Onkologiya-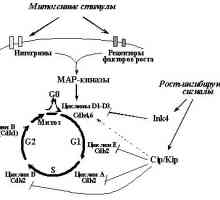 Onkologiya-
Onkologiya-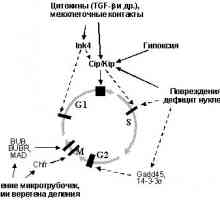 Onkologiya-
Onkologiya-- Onkologiya-
- Onkologiya-
- Otkrio sam genetski vibracijski urtikarije
 Genetika tumora
Genetika tumora Stanice koje utječu na matičnim stanicama. Generacijskih ciklus matičnih stanica
Stanice koje utječu na matičnim stanicama. Generacijskih ciklus matičnih stanica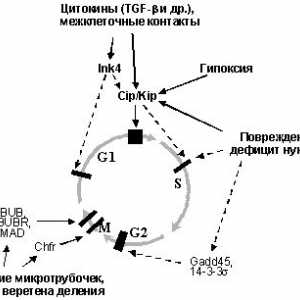 Onkologiya-
Onkologiya- Onkologiya-
Onkologiya-