Onkologiya-
B.P.Kopnin
Rak ruski istraživački centar. Blokhin RAMS, Moskva
izvor RosOncoWeb.Ru
01 0203 044. onkogeni, tumor supresori i poremećaja stanične morfogeneticheskihreaktsy
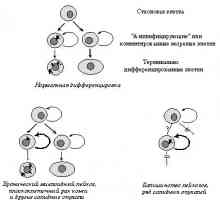
Svijetla osobina maligne stanice je"asocijalan" tip ponašanja koje je povezano na prvi ochereds oštećenja normalnih morfogenetskih reakcije - gubitak uzgoja kontaktnogotormozheniya, stjecanje sposobnosti proliferatsiinezavisimo vezanja na podlogu, mijenja adgezionnyhvzaimodeystvy, oblik i pokretljivost stanica, itd Te narusheniyavmeste s nekim drugim svojstvima, posebno sposobnostyusekretirovat proteolitičkih enzima i angiogenih čimbenika, rast predopredelyayutinvazivny znakova (prodiranja u okolna zdorovyetkani) i nakon toga metastaza (vtorichnyhochagov stvaranje tumora) [118]. Primarna uloga vozniknoveniiukazannyh gore poremećaja morfogenetski reakcije igrati izmeneniyafunktsii protooncogenes i / ili tumora prigušivač (Sl.7, 8).

Sl. 7. Promjene u aktivnosti proteina i potiskuju kontroliruemyhprotoonkogenami signalnih putova koje određuju vezanje ovisnost kletokot na podlogu i kontaktne inhibicije reproduktivnog (objašnjenje u tekstu)

Sl. 8. Podesiva Oncogene signalnih putova otvetstvennyeza mijenja morfologiju i fenotip lokomotorne neoplasticheskihkletok (objašnjenje u tekstu)
Svojstvena normalan stanični kontakt inhibicije razmnožavanja (prestanak širenja u dodir okruzhayuschimikletkami) odnose se prije svega na povećanu ekspresiju p16INK4a antitijelom i opuholevyhsupressorov p27KIP1 [37.119.120], da obuslovlivaetnedofosforilirovanie pRb i blokira ulazak u S-fazu [17] (točka 1). signalnog puta s plazma membrane kingibitoram ciklin-ovisne kinaze su nejasni. Samo što povećava ekspresiju u stanicama epitela E-kadherina zbog genske prijenosa, što dovodi do nakupljanja p27KIP1 iostanovke rasta stanica [121]. Nedavno, postoje dokazi o postojanju epitelnih stanica još jedan način blokira kletochnogotsikla kao odgovor na osnivanje stanica-stanica kontakata. Dokazano je da je formiranje sloja epitela uzrokuje nakupljanje p53, dok su mutacije E-kadherina i / ili disocijaciju stanica-stanica kontakata, naprotiv, uzrokuju destabizatsiyu p53 i, kao posljedica toga, utjecaj prekraschenieingibiruyuschego p21WAF1 na ciklina - CDK. Jer kod toga je jasno da se onkogeni potencijal E-kadherina mutacije odgovorne za razvoj želučanih i nasljednih oblika tumora raka mnogihdrugih [122] barem djelomično obuslovlenizmeneniyami regulacije staničnog ciklusa, apoptoze i kontrole geneticheskoystabilnosti [123].
Uz inaktivacije brojnih tumora potiskivača (E-kadherina, p53, p27KIP1, pRb), induciranih mutacija ili vezanje na virusnymionkobelkami (pRb - c E1A, E7, T-SV40- p53 - c E1B, E6, T-SV40-p27KIP1 - c E1A itd [7,10,124]), na gubitak kontakta može dovesti tormozheniyarazmnozheniya protooncogenes i hiperfunkcije, modifitsiruyuschihaktivnost signalni put pRb Cdk--E2F. Konkretno, mozhetbyt uzrokovano povećanom ekspresijom ili aktivacijom myc protoonkogenaRas (najprije uzrokuje degradaciju i p27KIP1 transaktivacija Cdc25a, drugi - p27KIP1 razgradnje i povećana ekspresija ciklina D1, vidi 1.).
Nezavisnost od vezanja ekstracelularnog matriksa (anchorageindependence). Preživjeti i razmnožiti, normalni tipovi kletkibolshinstva treba biti priključen na izvanstaničnog matriksu.Dva glavni čimbenici temelj ovog fenomena: nesposobnostrostovyh faktori aktivirati nevezan stanice kompleksytsiklin E - CDK2, koje su odgovorne za ulazak u S-fazi [125], te u mnogim induktsiyaapoptoza tipova stanica u odsutnosti adhezivnih interakcija (ova vrsta apoptoze ima poseban naziv "anoikis") [126]. I potiskivanje proliferacije i indukciju apoptoze u neprikreplennyhkletkah može biti povezana s aktivacijom p53 inducirane otkrepleniemkletok od podloge i odsutnosti signala od receptora integrina [58.127]. Za ulazni supresije u S-fazi aktiviranja dodavanje signalnogoputi od p53-p21WAF1, čini se da su odgovorni akkumulyatsiyap27KIP1 i uzorak se promatra u odsutnosti matrice kontaktovkletok [127,128]. Međutim, osim pokretanje kontrolne mehanizmovnegativnogo proliferacije (blokira ulaz u S-fazi je indukcija apoptoze), u odgovoru na odvojiti stanice iz matriksasuschestvuyut i nezavisnih mehanizama uzvodno reguliran proliferaciju preživljavanje stanica izazvanu vezanju integrina na belkamivnekletochnogo matriksa i naknadna aktiviranja neretseptornoytirozinkinazy FAK (fokalna adhezijska kinaza - tipka uchastnikaperedachi signali s integrinskim receptorima fizički vzaimodeystvuyuschegos citoplazmatske domene b-podjedinice integrina [118 , 126]) Najprije vezanju integrina na matriks i aktivacija FAK neobhodimydlya prolaz mitogeni signal receptorima za faktore rasta (EGF, PDGF), do konačnih MAP kinaza -. ERK1 / 2. (B neprikreplennoykletke signala iz ovih faktora rasta su blokirane na promezhutochnoyMAR kinaze MEK1, koji je zbog nekih razloga još nepoznatih fosforilaciju svojih ciljnih -. ERK1 / 2, [64]) drugo, aktiviruyaRas putem proteina adaptori Shc, integrin receptora ne tolkostimuliruyut mitogeno signala, nego i potiskuju anoikis (apoptoza), čini se da su zbog aktivacije Ras-signalni put PI3K-Akt / PKB [126] (vidi Sl. 2 odjeljak).
Ako pođemo od postojanja nekoliko mehanizama, opredelyayuschihzavisimost održivosti i / ili širenja stanica iz njihovog svyazyvaniyas matrice, postaje jasno da je za nastanak nezavisnosti harakternoydlya tumorskih stanica s prianjanjem vzaimodeystviyneobhodimo, navodno, nekoliko događaja koji, s jedne strane, omogućuje bi prevladati učinci supresor p53 (mutacija / deletsiietogo gena, prekomjernoj ekspresiji MDM2 onkogena et al.) i / ili p27KIP1 (mutacija / deletsii- prekomjerna ekspresija genima myc i ras, privodyaschayak degras kondenzacijom proteina, itd), a drugi -. bypass preryvaniemitogennogo signala na MEK1 kinaze (na primjer, zbog aktivatsiibelkov Src ili myc, uzrokuje aktivaciju ciklina E -Cdk2) i blok anoikis preko signalnog puta Ras-PI3K-PKB / Akt.
Promjene u obliku i staničnoj pokretljivosti. Promjenom oblika (morfologiju) - karakteristično svojstvo tumorskih stanica, što na mikroskopicheskomissledovanii dijagnozu maligne transformacije. Vosnove morfološke poremećaji su povezani izmeneniyatsitoskeleta, adhezivni interakcija stanica međusobno uc ekstracelularnog matriksa. Ukratko, oni su izražene u suprotnosti formirovaniyafokalnyh kontakata i pogoršanja vezanja stanica u matrici je neorganiziranost od aktin Microfilament sustava. To privoditk promjene u aktivnosti pseudopodia i pokreta znakova kletok.V općenito promatrano uzorak podsjeća na promjene nastale normalne stanice od djelovanja citokina motogenic - čimbenici koji stimuliraju kretanje stanica. Međutim, tzv lokomotornyyfenotip u maligne stanice su obično jako pretjerana, što omogućuje da se razlikuju morfološki od tumorskih stanica dvizhuscheysyanormalnoy stanica.
Molekularni mehanizmi koji stoje iza nastanka lokomotornogofenotipa kao u normalnim stanicama na motogenic poticaja učinio tijekom neoplastične transformacije, to je još uvijek daleko od jasno. Vyyavlenylish neke ključne čvorove preklapaju signalne krugove odgovorne za nastanak tih promjena. Kao što je poznato, mnogietsitokiny (npr HGF / SF, EGF, FGF, PDGF, IGF-1, itd.) I Yavlyayutsyaodnovremenno mitogenima motogenic i [118]. Tako, HGF / SF (HepatocyteGrowth Factor / Scatter faktor) je potentni mitogen za hepatocitima, a motogenic raznih epitelnim stanicama, posebno kletokmolochnoy žlijezda endotelne stanice i drugi. Motogenic učinak HGF / SFzavisit iz uzrokuje stimulaciju Ras-Raf-MAPK signalnim putovima : to se potvrđuje svojim ukidanja na inhibiciju signalnih MEK1 funkcije blokiranje ERK1 / 2 [129-131]. Na isti vremyaodnogo samo povećati aktivnost Raf, očito nedostatochnodlya rastavljanje u epitelnim stanicama E-kadherina i mezhkletochnyhkontaktov stimulacija pokretljivost stanica - Treba odnovremennayaaktivatsiya i PI3K, koji, kada HGF / SF mogu djelovati indutsirovatsyakak Ras-ovisna i Ras- strane nezavisni signalne staze [130,132]. Efektora PI3K, odgovorne za provedbu motogennogoeffekta nepoznato. Samo da ne PKB / Akt, ili Rac samipya sebi nije dao [130], iako se čini bazalna razina aktivnostiRac da će biti potrebni za provedbu pregrađivanje tsitoskeletai kontakt odvajanje stanica-stanica potrebnih za kretanje [132]. Također je pokazano da je jedan od važnih učinaka vyzyvaemyhHGF / SF stanica LMS je sniženje ekspressiigena fibronektina, koje mogu promijeniti interakciju prianjanja između matrice i kretanja [133]. Mutacije gena receptora HGF / SF-a (protoonkogenaMet) što dovodi do stalnog stimulaciju njegove tirozinkinaznoyaktivnosti su onkogeni mogu uzrokovati morfologicheskuyutransformatsiyu u kulturi stanica in vitro [134] i otvetstvennyza razvoj nasljednog papilarni karcinom bubrega kao i nekotoryhdrugih ljudskih bolesti (Tablica 1). , Molekularna osnova motogennyheffektov citokini proučili gore, ali se pretpostavlja da se, kao što je u slučaju HGF / SF-a, koji su povezani s stimulacijom Ras-MAPK-Ras signalizacije PI3Ki [135-139].
Konstitutivna ekspresija aktiviranog ras onkogena vyzyvaeti u fibroblastima i epitelne stanice i dramatičnog povećanja lokomotornoyaktivnosti trajne promjene karakteristične morfološke dlyaneoplasticheskih stanica [140.141] (bilješka, međutim, da ti izmeneniyahorosho očitovati samo u stanicama s nenormalnim p53 i / ili p16INK4a staza reguliruemyhsignalnyh - u drugim slučajevima, kao odgovor na Ras permanentnuyugiperekspressiyu mogu inducirati ili apoptoze ili ostanovkav G1, što smanjuje težinu Ras izazvane morfološka hizmeneny [117,142], pretpostavlja se da je nekoliko effektorovRas prvenstveno Raf obitelj GTPaza na Rho (Rac, cdc42, Rho) odgovorna za pojavu morfološke transformacije stimulacija lokomotorne aktivnosti neoplastičnih stanica [20,143] (sl. 8). Kao motogenic učinci citokina klyuchevuyurol ovdje, očito, igra aktiviranje dviju signalnih puteva :. Ras-PI3K-rac i Ras-Raf-ERK [20,143-145] Za transformaciju fibroblasta, očito dovoljno konstitutivna aktivacija bilo koje od tih putey.Deystvitelno, transdukcija aktiviranog Rac1, kao aktivni rovannyhRaf, Mos (kao što je Raf potiče MEK1), ili od MEK1 mozhetvyzyvat morfološki transformaciju glodavaca fibroblaste [146-150], i blokiranje u Ras-transformirane stanice ili funkcija MEK1 ili Rac, ali to dovodi do djelomičnog naslijeđena morfologicheskihizmeneny ali ne sprječava transformiranje [148,151]. Na zhevremya u epitelnim stanicama, pretvarajući dobro onkogenomRas aktivacije ERK-Raf-kaskada je dovoljna da uzrokuje morfologicheskuyutransformatsiyu [149]. Ekspresija aktivnog Rac vezani epitelialnysmkletkam znakove transformiranog fenotipa (kao pervuyuochered tvorbu lamellipodia, raffling membrane [143,152]), a snažno morfološki transformacija se postiže, čini se, samo kada su aktivirane u zajednički rac-ovisnog i Raf-ERK signalnyhputey [144.153].
O molekularnim događajima odgovornih za morfološke aktivaciju transformatsiyupri Rac i Raf-ERK signalnih puteva poznat vrlo malo.Stimulyatsiya MAP kinaze ERK1 / 2 vodi, kao što je poznato da aktivira ryadatranskriptsionnyh čimbenicima, osobito Elk1, fos, SRF. Umu da je jedan od najvažnijih posljedica hiperfunkcije Rac je stimulyatsiyadrugogo MAP kinaza kaskade (Slika 8.), Konačni produkt JNK kotorogo- - uzrokuje aktivaciju faktora transkripcije Jun i ATF2.Takim način i Raf-ERK- i rac-JNK-signalnih putova reguliruyutaktivnost AP-1 transkripcijski kompleksi (koji se sastoji od gomodimerovJun / lip heterodimera i to Jun / fos), čija je funkcija predstavlyaetsyavesma važno za izazivanje morfoloških promjena. Na etomsvidetelstvuyut kao sposobnost da uzrokuje onkogena lip transformatsiyukletok [154,155], i povrata transformiranih fenotipapri Jun suzbijanje funkcija [156.157]. Moguće je, međutim, chtotransformiruyuschy potencijal Jun (kao AP-1) nije povezan s izravnim deystviemna meta reguliraju adhezivne interakcije, organizatsiyutsitoskeleta i kretanje, te kako bi se dobilo autokrini proizvodnju petlje obuslovlennoystimulyatsiey faktora rasta / motogenic (EGF et al.), Koja se u njihovom red aktivira Ras i njegove efektore u drugim chastnostiRac obitelji GTP-aze Rho (cdc42, RhoA), morfologija promjene otvetstvennyeza [155,157] (Sl. 8). Pa se pretpostavlja da je formiranje lamclopodija pruža Rac i rafflinga, Cdc42- uzrokuje stvaranje filopodija i RhoA je uključen u formirovaniifokalnyh kontakata i stresnih vlakana [143.152].
Postojeći model obzir nastajanje lamellipodia kaksledstvie povećanja Rac induciranih aktivnosti protein Por1 [126.152] pojava stresnih vlakana - kao posljedica aktivatsiimiozinfosfatazy pod djelovanjem Rho-kinaze i formiranje fokalnyhkontaktov - kao rezultat vezanja na citoskeleta aktina i gelsolina belkovprofilina stimulira fosfatidilinozitolbifosfatom- jedan od Rho cilj [126.152]. Nejasno je, međutim, sve ove reakcije obespechivayutsyali signalizaciju isključivo između tsitoplazmaticheskimibelkami ili mijenjati set aktivnog transkripcije faktorovtakzhe pridonijeti tim procesima.
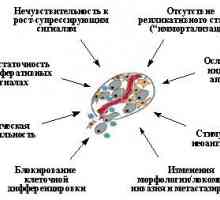
Dakle, dominantna uloga u stjecanju morfološke transformacije fenotipa lokomotornog igrati očito protooncogenes, promjene koje dovode do proteina aktivacije aktivnost semeystvaRas i / ili njegovog izvršnih - PI3K, Raf i eventualno RalGDS. Očigledno, samo ukupnost uzrokuju promjene u regulaciji aktivnostipsevdopody, montaže / demontaže citoskeleta i žarišne kontaktovpri simultano promjene u aktivnosti velikom skupu transkriptsionnyhfaktorov daje na kraju kupiti taknazyvaemogo stanice "potpuno preobrazila" fenotip, koji određuje agresivnu prirodu rasta. Pri tome treba imati na umu da je za pojavu tih promjena zahtijeva inaktivacije opuholevyhsupressorov (p53 i / ili p19ARF p16INK4a), zaštitna organizmot pojavu njemu klonova stanica s konstitutivno aktivirovannymiRas-MAP-kinaza signalnim putovima. Osim toga, pretvaranje vyrazhennostmorfologicheskoy kretanje i sposobnost povećanja ekspresije umenshayutsyapri neki drugi tumora supressorov- E-kadherin [118] i pRb [158]. Osim toga, druge dvije neoplastična stanica vazhneyshihpriznaka - ublažavanje tormozheniyarazmnozheniya kontakt i kupiti od neovisnosti supstrata - zavisyatv prvenstveno na inaktivacije određenih tumora potiskivača (p53, p27KIP, Rb, E-kadherina). Dakle, čini se očigledan chtodlya izgled karakterističan za tumorske stanice izmeneniymorfogeneticheskih reakcije zahtijevaju više genetičkih događaja (mutacije) koji utječu i tumorskih smetnji i proto-onkogena.
5. onkogena i tumor supresori u neoangiogenezomNeoangiogenezom - stvaranje mreže kapilara endotelialnyhkletok obloge male venule - preduvjet za tumor nodula dalneyshegorosta, dosegao promjer 2-4 mm [118.159] .Priobretenie sposobnost stimulirovatproliferatsiyu neoplastične stanice i migraciju endotelnih stanica bio je spojen s dva glavna događaja: prestanak izlučivanja čimbenika koji inhibiraju angiogenezu (trombospondin et al.), te su citokini uvelicheniemproduktsii faktori rasta i motogenic dlyaendoteliotsitov (iznad oko izmjenične VEGF i FGF, EGF, TGF-a), praćeno povećanim izlučivanjem i / ili aktivnost proteaza djeluju proteolize ekstracelularnog matriksa i invaziji tkiva endoteliotsitovv neoplazme.
Ključni ulogu u nastanku fenotipa angiogeneze neoplasticheskihkletok igra očito tumora supressorar53 inaktivacije funkciju koja kontrolira ekspresiju nekoliko inhibitora i stimulyatorovangiogeneza. Tako, geni trombospondin 1 i 2 je mishenyutransaktivatsionnogo koraka p53 [160,161] i p53 transkpitsiyu genaVEGF, s druge strane, inhibira [162,163]. Zajedno sa aktiviranim sposobnostyur53 u odgovoru na hipoksiju [164], ovaj predopredelyaetesche mehanizam kojim normalno funktsionirovanier53 može zaštititi protiv rasta tumora: hipoksija, voznikayuschayav centar neoplastičnih kvržica inducira p53 i kako sledstvieapoptoz ili zastoj staničnog ciklusa, uz povysheniemsekretsii trombospondin i smanjenje ekspresije VEGF, što dolzhnopredotvraschat neovaskularizaciju čvor. Naravno, zbog toga, chtoinaktivatsiya p53 može biti važan korak u stjecanju sposobnostistimulirovat angiogeneze. Doista, analiza mehanizmovvozniknoveniya angiogenetski fenotip u ljudskoj fibroblastahbylo otkrili da je u većini slučajeva inaktivatsiyar53 je početni događaj [165].
Kako bi se dodatno poboljšala sposobnost stimuliranja angiogenezu ekspresiju mozhetprivodit onkogena, naročito semeystvaRAS onkogeni, uzrokujući transkripcijsku aktivaciju AP-1 i, kao posljedica toga, s jedne strane, povećana sekrecija VEGF, kotorogosoderzhit gena odgovarajućim elementima za AP-1 [165-169], i s druge strane, - je broj povećan proizvodnju matriks metaloproteinaze (MMP-9 / kolllagenazaIV, MMP-1, itd.) [169-171] čiji se geni također regulirana AP-1 [172.173] i druge Ras-inducirajućih transkripcijskih čimbenika, osobito Ets1 [172,173].
Treba primijetiti da je slijed događaja privodyaschihk razvoj angiogenetski fenotip može biti različit: u fibrosarkomahu transgenskih miševa izgled angiogenetski fenotip initsiirovalosne mutacije p53 i povećana ekspresija JunB i c-Jun komponentovkompleksa AP-1, a što dovodi do povećanja izlučivanja opuholevyhkletok FGFb [174] , Podaci o ulozi onkogena i drugih iopuholevyh smetnji u regulaciji angiogeneze. Tako, Myc podavlyaettranskriptsiyu trombospondin 1 [26], a tumorske supressorVHL, mutacija koje uzrokuju sindrom von Hippel Lindau (razvitiemnozhestvennyh hemangiomi) i karcinom bubrega [175] (Tablica 2), osuschestvlyaetnegativnuyu regulacija VEGF ekspresije gena u strome kletkahgemangiom [176] bubrežne i epitelne stanice [177]. Dakle, očito je da su promjene u aktivnosti određenih tumora supressorovi onkogena igrati ključnu ulogu u poticanju angiogeneze.
6. Uloga onkogena i tumorskih potiskivača stjecanje sposobnostik metastaziranjeMetastaza - formiranje sekundarne žarišta tumora rosta- najopasniji manifestaciju progresije tumora, yavlyayuscheesyaosnovnoy uzrok smrti u pacijenata oboljelih od raka. Kako bi metastaza, stanica mora steći brojna svojstva: sposobnost da prodre u glubinuokruzhayuschih normalnim tkivima (uključujući krv ili limfaticheskiesosudy), sposobnost da preživi nakon što je pogođen u posudama i zatempenetrirovat ih i razmnožiti u neobično za ovaj tipakletok mikro okoliš, daje novi fokus rast tumora [118] .tak, sposobnost metastaziranja nastaje izkompleksa jednostavnije mogućnosti velikog dijela lokomotornog which- stjecanje fenotipom povećana proteolitičkih aktivnost je sposobnost stimuliranja angiogenezu i stvoriti za bijeg tumorske stanice temsamym od primarnog tumora, vozniknovenienezavisimosti iz supstrata i suzbijanje apoptoze - već rassmotrenav prethodnom odlomku. Pojava svake od ovih znakova uvelichivaetveroyatnost razvoj metastatskog potencijala. Međutim naiboleevazhnymi zastupljeni gena i proteina, kao što su p53, Ras i Src, promjenama koje rezultiraju vozniknoveniyusrazu aktivnosti nekoliko komponenata metastatskim fenotipom, kao i, Krometogo - genetičkoj nestabilnosti koja olakšava izbijanja dopolnitelnyhpriznakov potreban za metastaze. Zanimljivo, p53, što znatno povećava disfunkcije sposobnostkletok metastazirati na modelnim sustavima in vivo [58.178.179], između ostalog izravno aktivira transkripciju nedavno otkrytogogena KAI1 [180] - transmembranski protein koji tvori kompleksys E-kadherin. Gubitak ekspresije proteina KAI1 zbog vtom uključujući inaktivacije p53 [180], naravno obnaruzhivaetsyav različitih humanih tumora u kasnim stadijima bolesti (60-90% od raka prostate, gušterače i mliječnih žlijezda, melkokletochnogoraka pluća, rak hepatocelularnog, itd.) [181- 185], a ekspresija vosstanovlenieego uzrokuje inhibiciju metastatskog postupka [186-188]. Kalcij-vezujući protein S100A4 / MTS1 / CAPL (metastazin), naprotiv, prekomjerno eksprimiran u kasnim stadijima ljudskog razvoja razlichnyhnovoobrazovany [189], a ima sposobnost napada i pridavatkletkam metastaziranja [190-192]. Egoekspressiya također uzrokuje pleiotropskim efektima: vodi kumensheniyu sadržaja E-kadherina [193], inhibiciju sinteze ingibitorametalloproteinaz TIMP-1 [194], promjene u dinamici regulacije reorganizatsiytsitoskeleta kao rezultat inhibicije fosforilacije tyazheloytsepi miozina [195] i, eventualno, za odvajanje i funktsionalnoyinaktivatsii p53 [ 189].
Naravno, promjene gore opisani nisu iscrpni perechenkontroliruemyh tumora smetnji i / ili protoonkogenamipriznakov da mogu igrati značajnu ulogu u priobreteniisposobnosti da metastaziraju. Dakle, znatan interes predstavlyaetopisanny G.I.Deychman takozvani "H2O2CA + PGES"fenotip se sastoji u povećanju antioksidativno djelovanje sekretsiiprostaglandina E2 i kao rezultat toga, čak iu bolsheyustoychivosti stjecanja faktora prirodnog imuniteta stečena antitumorsko otpor [196.197]. Taj fenotip indutsiruetsyav uzgojene stanice in vitro, transdukcijom opredelennyhizoform onkogena v-src, ali ne i druge istraživani geni (aktivirovannyyH-ras, myc, bcl-2, mutantni p53, E1A, LT SV40), ili nastaje vprotsesse rast tumora in vivo, naznačen time što je metastatski sposobnostkletok korelira s to. Očito, u sljedećih nekoliko godina čekamo drugih važnih spoznaja koje bacaju svjetlo na mehanizme metastazirovaniyai ulogu u ovim procesima tumora suppressors i onkogena.
 Maligni insuloma. Morfologija tumora Langerhansovih otočića
Maligni insuloma. Morfologija tumora Langerhansovih otočića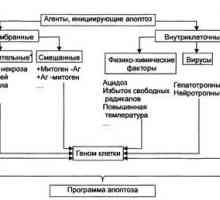 Apoptoza. Malignosti stanica tijela
Apoptoza. Malignosti stanica tijela Rak se širi uz pomoć „lošeg” kolesterola
Rak se širi uz pomoć „lošeg” kolesterola Diferencirani adenokarcinom želuca
Diferencirani adenokarcinom želuca Klasifikacija raka (tumora), gušterače ICD 10
Klasifikacija raka (tumora), gušterače ICD 10- Liječenje zračenjem za rak pluća
- Najviše predstavlja adenoma (90%). tumor karakterizacija malignog ne temelji na podacima…
- Zdravlje Enciklopedija, bolest, lijek, liječnik, apoteka, infekcija, sažetak, seks, ginekologije,…
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
 Onkologiya-
Onkologiya-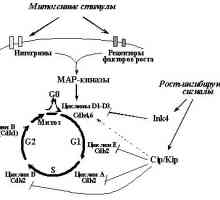 Onkologiya-
Onkologiya-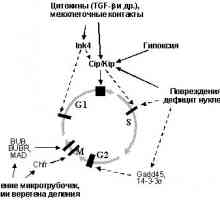 Onkologiya-
Onkologiya-- Onkologiya-
 Onkologiya-
Onkologiya-- Onkologiya-
- Onkologiya-
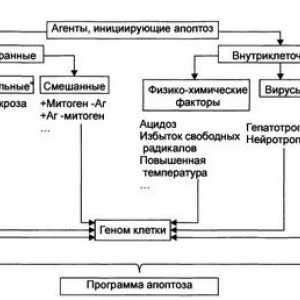 Apoptoza. Malignosti stanica tijela
Apoptoza. Malignosti stanica tijela Klasifikacija raka (tumora), gušterače ICD 10
Klasifikacija raka (tumora), gušterače ICD 10 Genetika tumora
Genetika tumora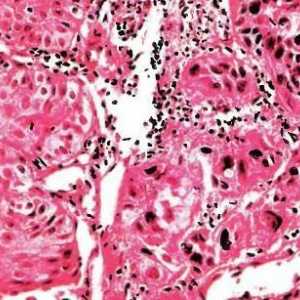 Diferencirani adenokarcinom želuca
Diferencirani adenokarcinom želuca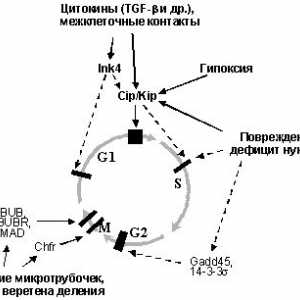 Onkologiya-
Onkologiya-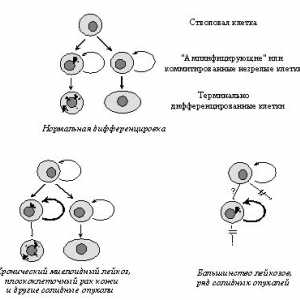 Onkologiya-
Onkologiya-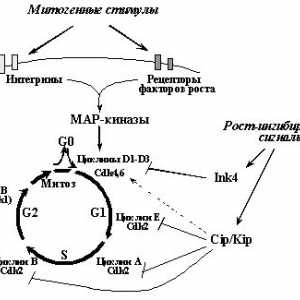 Onkologiya-
Onkologiya-